

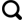
0 引言
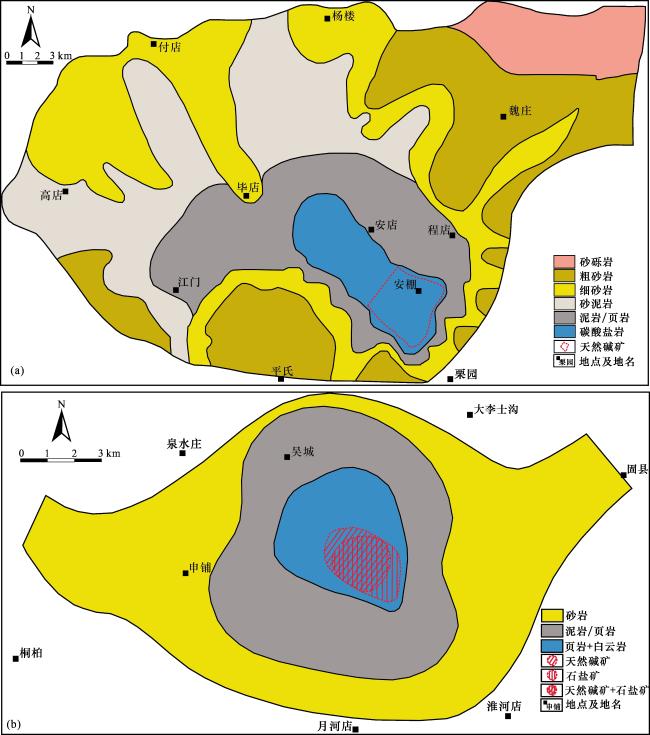
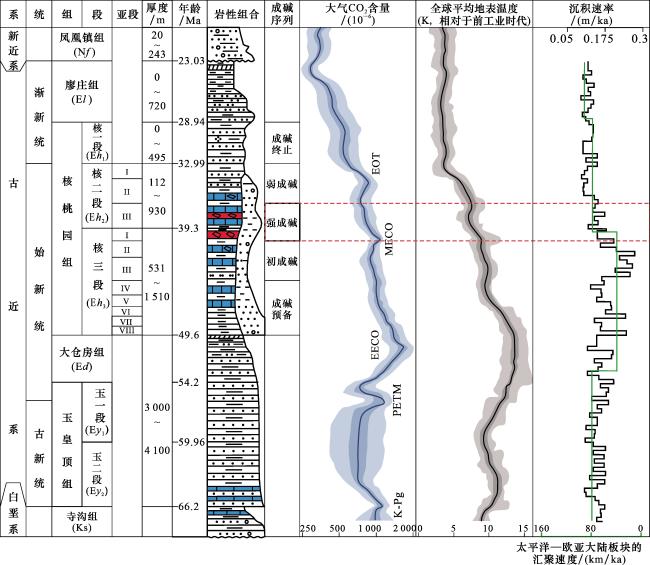
碱湖(碳酸盐型湖泊)以高碱性水体(pH>9)及以CO 、HCO 为主的阴离子组成为特征,显著区别于以CO —SO —Cl-组合为主的硫酸盐型湖泊和以Cl-为主的氯化物型湖泊[1]。尽管地质历史时期发育的碱湖相对较少,但却是油气、天然碱矿和硼矿的重要地质载体[2-4]。例如准噶尔盆地下二叠统风城组碱湖烃源岩,是盆地西北缘百里油区形成的物质基础[5];世界上最大的天然碱矿床——美国西部绿河组(Green River Formation)天然碱矿,也形成于古近纪始新世碱湖环境[6]。此外,古代碱湖的极端碱性水体条件孕育了独特的生物群落,使其成为研究深时极端古生态的关键档案[7]。古代碱湖沉积序列中,普遍发育天然碱矿层—白云岩—富有机质泥页岩(或油页岩)的特征性共生组合,例如美国大绿河盆地(Greater Green River Basin)绿河组的天然碱矿层,顶底板多由油页岩和白云岩组成[8];我国规模最大的古代天然碱矿——河南安棚与吴城天然碱矿,其矿层厚度中心与富有机质泥页岩和白云岩的沉积中心高度重合(图1),在垂向上三者也常呈薄、中厚层状紧密互层,构成韵律性沉积组合(图2)[9-10]。
古代碱湖特征性的天然碱矿层—白云岩—富有机质泥页岩(或油页岩)共生组合,揭示天然碱—白云石—微生物之间存在成因关联。这种成因联系的核心机制为“微生物群落—白云石互馈作用”,包含以下3个层次:①碱湖微生物促进白云石沉淀:在碱湖极端碱性(pH>9)水体中,蓝细菌、硫酸盐还原菌和产甲烷菌等嗜碱/耐碱微生物主导生态系统[7]。已有研究证实,这些微生物可通过改变胞外微环境(如提升局部pH值、消耗SO 、产生CO 等),以及提供初始成核位点(如胞外聚合物EPS、细胞膜等)等方式,克服白云石形成的动力学屏障,促进白云石沉淀[11]。②白云石沉淀驱动微生物群落演替:白云石沉淀会固定碱湖水体中的Mg2+/Ca2+,并通过消耗CO 影响CO —HCO 缓冲系统,引发水体离子浓度梯度和pH波动,改变微生物群落外部环境,进而驱动微生物群落演替[12]。③互馈终止与天然碱矿物形成:碱湖微生物群落—白云石互馈作用持续进行导致水体中的Mg2+/Ca2+/ SO 被逐步消耗,当其中一种或几种离子浓度降至临界阈值以下,互馈循环终止。此时,碱湖高碱度、高Na+和CO 的水化学条件,为天然碱矿物的形成提供了物质基础,随着水体进一步蒸发浓缩,天然碱矿物便开始沉淀析出[6,13]。
综上,碱湖微生物群落—白云石互馈作用不仅控制天然碱矿物的形成,而且对有机质富集有显著影响(如微生物群落演替影响着古生产力)。深入揭示该互馈机制及其对有机质富集的影响,既可指导古代碱湖相关矿产资源勘探,也是对深时极端环境古生态研究的有益补充。
1 碱湖水化学演化与生物群落结构
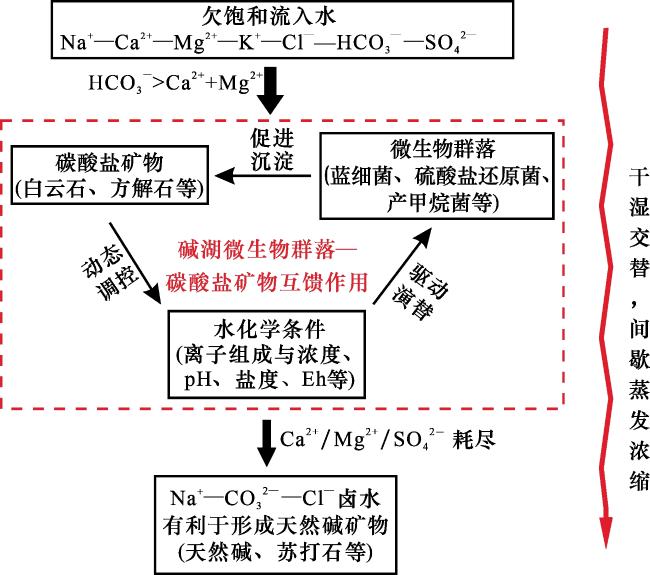
在持续蒸发浓缩条件下,碱湖水化学演化通常遵循特定的序列[1](图3路径①)。初始离子欠饱和水体(Na+—Ca2+—Mg2+—K+—Cl-—HCO —SO 体系)汇入湖盆后,随着蒸发作用增强,碳酸盐矿物优先析出,此时会出现“碳酸盐矿物分异”现象。碱湖HCO 浓度显著高于Ca2+和Mg2+,使得Ca2+与Mg2+率先消耗殆尽,故形成富Na+—CO —SO —Cl-的碱性卤水(如美国加州莫诺湖)。由于碱湖中的CO 和Mg2+浓度较高,且高pH条件能够降低Mg2+的水合作用,并抑制方解石等竞争矿物的形成,因此碱湖沉积物中的白云石丰度通常高于方解石、文石等其他碳酸盐矿物[14-16]。进一步蒸发浓缩过程中,硫酸盐还原菌(SRB)等微生物通过代谢作用将SO 还原为S2-,促使水化学条件转变为Na+—CO —Cl-主导(如东非马加迪湖)。当此类卤水继续蒸发浓缩,水体活度积超过天然碱矿物的溶度积时,天然碱矿物便开始沉淀析出。
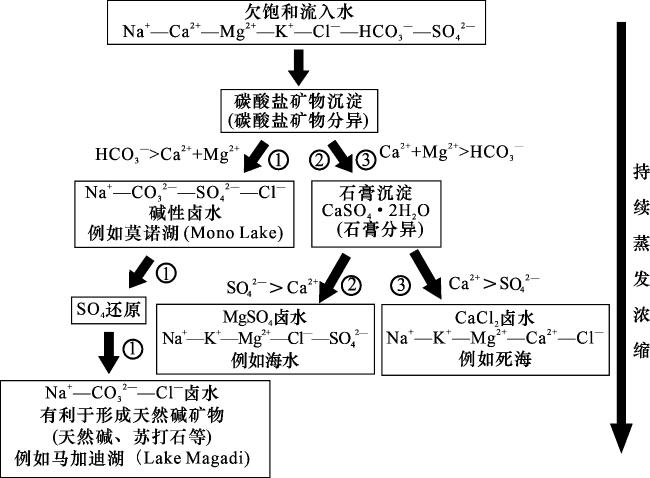
相较之下,硫酸盐型与氯化物型湖泊在持续蒸发浓缩过程中的水化学演化路径与碱湖(碳酸盐型湖泊)存在显著差异(图3)。其核心控制因素在于不同类型湖泊初始离子组成的差别,硫酸盐型和氯化物型湖泊的HCO3 -浓度常低于Ca2+和Mg2+,导致碳酸盐矿物沉淀阶段HCO 被率先耗尽,继而触发石膏沉淀。此后演化路径取决于SO 与Ca2+的相对含量:当SO >Ca2+时,石膏沉淀使Ca2+优先耗尽,形成富Na+—K+—Mg2+—Cl —SO 卤水,最终沉淀MgSO4等硫酸盐矿物(图3路径②)。而当Ca2+>SO 时,石膏沉淀将使SO 率先被耗尽,卤水将进一步演化为Na+—K+—Mg2+—Ca2+—Cl-体系,后期以石盐等盐类矿物的析出为终点(图3路径③)。
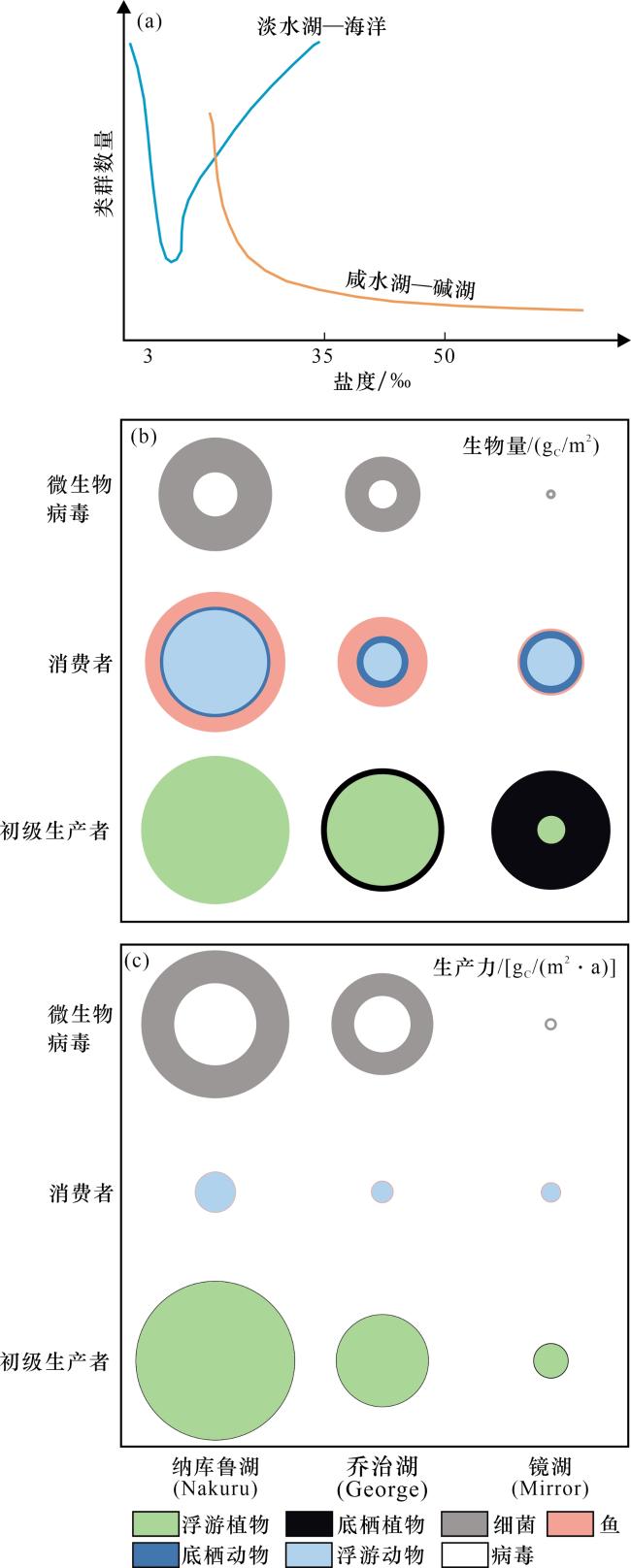
图4 湖泊水化学条件对生物类群数量、生物量和生产力的影响[7](a)淡水湖—海洋和咸水湖—碱湖生物类群数量随盐度变化的趋势;(b)碱湖(肯尼亚纳库鲁湖)、热带富营养湖(乌干达乔治湖)和温带中营养湖(美国镜湖)的生物量对比;(c)碱湖(肯尼亚纳库鲁湖)、热带富营养湖(乌干达乔治湖)和温带中营养湖(美国镜湖)的生产力对比。注:圆面积代表不同生物类型的相对贡献,细菌生产力根据生物量估算得出;3个湖泊的总生物量分别为21.3 gc/m2, 11.3 gc/m2和7.4 gc/m2,总生产力分别为1 900 gc/(m2·a), 740 gc/(m2·a)和65 gc/(m2·a) Fig.4 Effects of lake water chemistry on the number of taxa, biomass and productivity[7] |
2 碱湖微生物群落与白云石的互馈作用
最早在潟湖环境中微生物成因原生白云石的理论取得突破,并在现代、古代盐(碱)湖以及淡水—微咸水湖得到进一步发展[20-22]。目前已发现硫酸盐还原菌、产甲烷菌、蓝细菌和胞外聚合物(EPS)等微生物(有机物)可通过改变细胞周围微环境、提供初始成核位点等方式克服动力学屏障,促使白云石形成[23-25]。尽管近年来被证实一些无机物,例如溶解NH3和SiO2,也能在实验室低温条件下促进白云石形成[26-27],但微生物(有机物)仍被认为是低温条件下促使白云石形成的关键媒介[11]。碱湖的极端碱性环境为蓝细菌、硫酸盐还原菌和产甲烷菌等嗜碱/耐碱微生物主导的生态系统,它们可通过提高水体pH值和CO 浓度、降低SO 浓度,并导致细胞带负电荷,使Mg2+/Ca2+等金属阳离子易被吸附到微生物细胞膜和胞外聚合物(EPS)上,克服白云石形成的动力学屏障,促进原生白云石的形成[28](图5)。但它们之间的区别在于,不同种类的微生物生存于碱湖不同的垂直分带内,其生存条件中氧气、SO 浓度等因素存在差异(图6)。微生物成因原生白云石在现代和地质历史时期的碱湖都普遍发育,例如乌鲁木齐红雁池水库剖面中二叠统芦草沟组的湖相微晶白云岩中,包含纹层状的微—纳米级卵状和椭球状白云石,它们的晶体形态、显微结构和碳同位素比值共同指示其形成与产甲烷菌等微生物有关[29]。此外,玛湖凹陷二叠系风城组和泌阳凹陷安棚地区古近系核桃园组碱湖沉积序列中也发现了微生物成因原生白云石[4,30]。
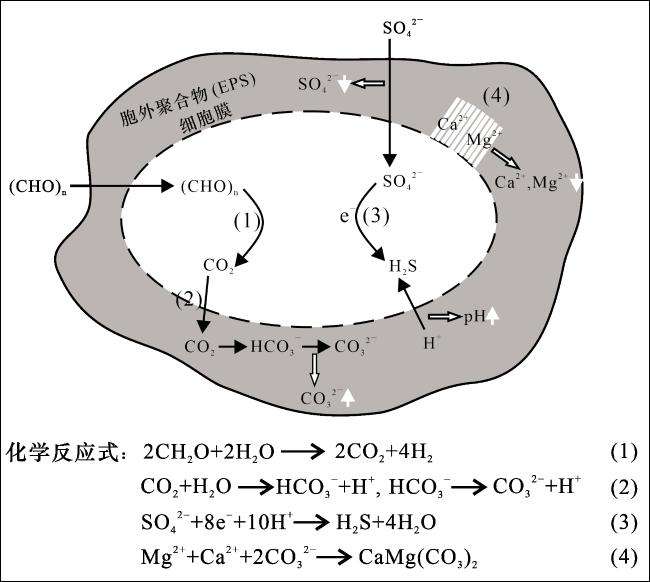

从表层向下随着溶解氧浓度的降低,碱湖可依次划分出氧化带、贫氧带、厌氧—硫化带以及硫酸盐—甲烷转换带(图6)。垂向上各分带内,受控于溶解氧浓度、pH、盐度、光照等水体条件变化,微生物类型与代谢功能也随之变化[31]。例如在氧化带内,蓝细菌等自养需氧微生物利用氧气分解有机质,形成HCO 、H+等;在厌氧—硫化带内,硫酸盐还原菌将SO 还原为S ,并产生HCO3 -;而在硫酸盐—甲烷转换带内,硫酸盐还原菌与异养产甲烷菌共同作用,还原SO 与CH4循环的同时,生成HCO [32-33]。以上微生物活动所产生的HCO 为碱湖原生白云石沉淀提供了充足的物质来源。与此同时,碱湖白云石沉淀固定Mg2+/Ca2+,消耗CO3 2-影响CO —HCO 缓冲系统,调控水体离子组成与浓度,进而改变微生物群落外部环境。前已述及,碱湖的极端碱性条件对微生物群落结构的影响远超种间相互作用,因此群落外部环境的改变将会驱动微生物群落演替[12,18]。综上,碱湖微生物群落通过改变胞外微环境、提供初始成核位点等方式促进白云石沉淀;而白云石沉淀通过影响水化学条件,改变微生物群落外部环境,驱动微生物群落演替。至此形成完整循环:微生物群落活动—促进白云石沉淀—改变水化学条件—驱动微生物群落演替—新微生物群落活动,此即碱湖“微生物群落—白云石互馈作用”。
在持续蒸发浓缩的条件下,碱湖水化学演化会遵循从初始离子欠饱和水(Na —Ca —Mg —K —Cl-—HCO —SO ),到Na —CO —SO —Cl-,再到Na —CO —Cl-型卤水的过程(图3)。然而,碱湖演化过程中气候通常不是持续干旱,而是呈现出干旱背景下的频繁干湿交替特征[34]。在此背景下,碱湖的湖平面会出现高频振荡[7],在此过程中水化学条件演化并非如图3路径①所示的线性过程,而应如图7所示,在HCO 浓度显著高于Ca2+和Mg2+的条件下,出现“微生物群落—白云石互馈作用”,在此过程中白云石沉淀逐步消耗Mg2+/Ca2+,硫酸盐还原作用使水体的SO4 2-浓度逐渐降低,当Mg2+/Ca2+/SO 中的一种或几种离子浓度降至临界阈值以下时,互馈循环即终止。此时,碱湖为Na —CO —Cl-型卤水主导,当气候继续向干旱演变,天然碱矿物便开始沉淀析出。
3 碱湖有机质富集机理
陆相湖盆沉积有机质富集受控于有机质生产、有机质保存和有机质稀释3个方面要素[35]。碱湖通常在干旱气候背景下形成,陆缘碎屑输入较少,因此相比于有机质生产和有机质保存,有机质稀释(主要受控于沉积速率)对有机质富集的影响相对较弱。
3.1 碱湖有机质生产
表1 现代不同类型生态系统净初级生产力对比[7]Table 1 Comparison of net primary productivity of different type of modern ecosystems[7] |
| 生态系统 | 净初级生产力/[ ] | 生态系统 | 净初级生产力/[ ] |
|---|---|---|---|
| 北极苔原* | 445 | 25个温带湖泊(25°~60° N) | <2 000 |
| 针叶林* | 2 840 | 热带潟湖(墨西哥湾) | 4 450 |
| 阔叶林* | 3 547 | 热带海草草甸(印尼) | 4 177 |
| 热带潮湿森林* | 6 317 | 热带淡水湖泊(Tana湖) | 887 |
| 温带河口(切萨皮克湾) | 1 666 | 碱湖(Elementaita湖) | 4 282 |
| 易发洪水的河流(瑞士) | 1 825 | 碱湖(Bogoria湖) | 4 875 |
| 瓦登海(德国) | 469 | 碱湖(Nakuru湖) | 6 854 |
|
碱湖极高的净初级生产力与其独特的高pH水化学条件密切相关。碱湖可以从大气中吸收更多的CO2,提升湖水中溶解无机碳(DIC)的含量,同时使生物的光合作用不受CO2浓度限制,促使能进行光合作用的生物量增加[36]。由于P和N通常是限制湖泊初级生产力的重要营养元素,而碱性溶液可以增加磷酸盐和硝酸盐的溶解度,进而极大促进碱湖中生物的生长繁殖[37]。碱湖中的游离硫化物主要以HS-状态存在,鉴于其对生物的毒性要远低于H2S和多硫化物,因此游离硫化物对碱湖生物的破坏较小,间接促进生物繁盛[38]。当pH高于9时,硅质溶解度会随pH值升高呈指数增长,因此碱湖中溶解硅酸盐含量较高,有利于硅藻勃发[39]。综上,碱湖独特的高pH水化学条件可活化营养元素,中和有害物质,使初级生产者繁盛,进而提高初级生产力。此外,碱湖的极端碱性条件使其生物类群较少,极短的食物链,也是造成其高剩余初级生产力的一个重要原因。
3.2 碱湖有机质保存
缺氧还原环境对有机质的保存更为有利,大型湖泊温跃层和化跃层以下的稳定缺氧水体通常是有机质保存的最理想场所。但碱湖在蒸发浓缩背景下形成,水体深度较小,例如肯尼亚纳库鲁(Nakuru)碱湖在1930—2000年,最高水位仅不足4.5 m,且通常在1~3 m之间波动[7]。然而,碱湖沉积物的有机质丰度一般却较高,例如准噶尔盆地二叠系风城组碱湖烃源岩的有机碳含量主要集中在1.0%~2.0%之间,最高可达3.8%[3];泌阳凹陷古近系核桃园组碱湖烃源岩的有机碳含量主要介于1.5%~3.0%之间,最高可达5.2%[40]。据此,李长志等[37]提出早期硅化和热泉输入是碱湖有机质在浅水环境下有效保存的重要机制。其中早期硅化是指碱湖的富有机质层被硅质覆盖,有效阻止其进一步降解;而热泉输入是指当气候干旱期碱湖水位过低时,不受气候影响的地下热泉亦为湖泊输入水量,减少有机质暴露降解的概率,促进有机质保存。以上2种碱湖有机质保存机制均已在现代(如肯尼亚博戈里亚碱湖)和古代(如玛湖凹陷二叠系风城组)碱湖中被证实[37,41]。而且与高碱度伴生的高盐度会降低水体溶解氧含量,因为盐离子会竞争水分子与氧分子间的结合位点,从而降低氧分子的溶解量。
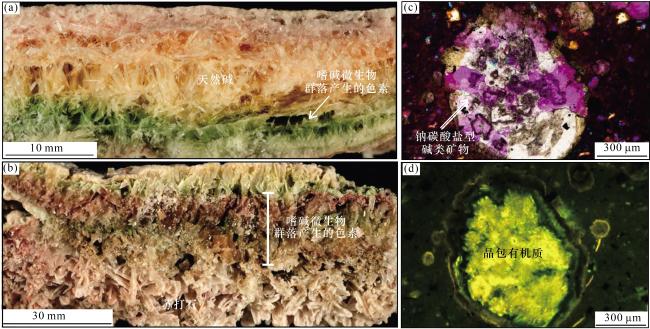
在此基础上,本文补充提出天然碱矿物壳的保护也是碱湖有机质有效保存的另一个重要机制。在干湿交替,间歇蒸发浓缩的背景下,碱湖存在微生物群落—白云石的互馈作用,互馈作用会使得Mg2+/Ca2+/SO 被逐步耗尽,形成有利于天然碱矿物形成的卤水,并随着进一步蒸发浓缩,开始形成天然碱矿物。而这些在同生—准同生或早期成岩作用阶段形成的天然碱矿物会包裹在有机质外层,形成天然碱矿物保护壳。例如东非纳西克安格达碱湖(Nasikie Engida)中嗜碱微生物群落产生的色素会被天然碱、苏打石外壳包裹[6][图8(a),图8(b)],而准噶尔盆地二叠系风城组烃源岩中也发现了包裹着钠碳酸盐型碱类矿物的晶包有机质[3][图8(c),图8(d)]。以上实例证实天然碱矿物保护壳是碱湖有机质有效保存的另一重要机制。一方面,在同生—准同生阶段形成的天然碱矿物壳会使得有机质快速通过水柱沉淀至水—沉积物界面,从而减少沉降过程中有机质所遭受的分解破坏。另一方面,在早期成岩作用阶段天然碱矿物壳能起到隔绝氧气、减少微生物分解的作用,进一步保护其中的有机质。
综上,碱湖独特的水化学条件使其能够活化营养元素,中和有害物质,提高初级生产者占比,形成异常高的初级生产力;同时,可通过早期硅化、热泉输入和形成天然碱矿物外壳减少有机质的分解,对浅水有机质进行有效保存。因此,碱湖是沉积有机质富集的理想场所,也是发育具有独特生烃行为优质烃源岩的特殊环境[42]。
4 泌阳凹陷古近纪碱湖微生物群落—白云石互馈作用
泌阳凹陷位于南襄盆地东部,面积约为1 000 km2。其东南部安棚地区赋存我国规模最大的古近系天然碱矿床,矿体分布面积约为50 km2。安棚天然碱矿主要发育在古近系始新统核桃园组三段顶部和二段底部,共计17层,呈层状—似层状展布,平面投影呈不规则圆形,埋深在1 477~2 552 m之间,累计厚度为1 500 m[9]。安棚天然碱矿层在空间上与白云岩和富有机质泥页岩紧密共生[图1(a),图2(a)]。矿石类型以苏打石(NaHCO3)为主,其次为天然碱(Na2CO3·NaHCO3·2H2O)和碳氢钠石(Na2CO3·3NaHCO3)[43]。矿物相平衡实验表明苏打石形成于高CO2浓度、广温条件下;天然碱形成于高温、广CO2浓度条件下;而泡碱则形成于低温、低CO2浓度条件下[44]。因此,安棚以苏打石为主的天然碱矿床指示其发育时期的碱湖水体具有高CO2浓度特征。
微形貌特征和显微结构是微生物成因原生白云石判识的最重要也是最常用的标志[29]。泌阳凹陷安棚地区核三段上—核二段下含碱层系中,发育天然碱矿层、白云岩和富有机质泥页岩的共生组合。其中白云岩主要由泥晶白云石构成,可见破碎的钙质生物碎屑[图10(a)];富有机质泥页岩中常见白云石纹层,与黏土和有机质组成的纹层互层[图10(b)]。扫描电镜下,白云石主要为微纳米级球状和杆状,可见由纳米微粒聚集黏结而成的纳米质点结构,能谱结果显示白云石Ca含量较高,Fe含量较低[图10(c),图10(d)]。结合含碱层系白云岩的有序度介于0.3~0.5之间,以及含碱层系的镜质体反射率通常在0.7%以下,综合表明泌阳凹陷安棚地区核三段上—核二段下的含碱层系中发育微生物成因的原生白云石。

5 结论
(1)碱湖的极端碱性(pH>9)水体条件使蓝细菌、硫酸盐还原菌和产甲烷菌等嗜碱/耐碱微生物主导生态系统,它们可通过改变胞外微环境、提供初始成核位点等方式,促进白云石形成。而白云石沉淀固定Mg2+/Ca2+,引发离子梯度与pH波动,改变碱湖水化学条件,驱动微生物群落演替,形成微生物群落—白云石互馈作用。
(2)在干湿交替气候,水体间歇蒸发浓缩背景下,碱湖的湖平面出现高频振荡,微生物群落—白云石互馈作用逐步消耗Mg2+/Ca2+/SO ,当离子浓度降至临界阈值以下时,互馈循环终止。此时碱湖为Na —CO —Cl-型卤水,随着水体进一步蒸发浓缩,天然碱矿物开始沉淀析出,形成古代碱湖特征性的天然碱矿层—白云岩—富有机质泥页岩共生组合。
(3)碱湖独特的高pH水化学条件可活化营养元素,中和有害物质,提高初级生产者占比,形成极高的初级生产力;同时早期硅化、热泉输入和天然碱矿物外壳的形成可减少碱湖有机质的分解,二者共同促进碱湖沉积物(岩)中有机质富集,为碱湖优质烃源岩发育提供物质基础。
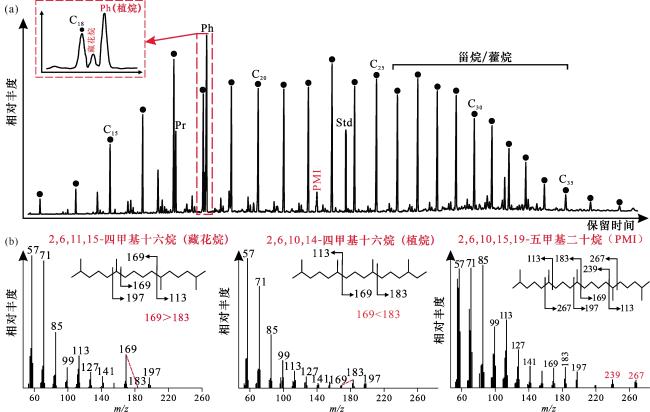
(4)泌阳凹陷安棚地区古近系核三段顶—核二段底含碱层系的矿石类型主要为苏打石,大气、岩浆深源和微生物活动共同为碱湖提供CO2。该层段发育球状/杆状、纳米质点结构的原生白云石,且检测出藏花烷、PMI等特征生物标志物,为古代碱湖微生物群落—白云石互馈作用提供直接证据。
 甘公网安备 62010202000678号
甘公网安备 62010202000678号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}