

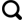
0 引言
烃源岩的形成取决于成烃母质生物的生存环境和有机质良好的保存条件,这两大因素从根本上来说又取决于沉积时期古生产力、营养物质供应、古气候及古海洋水体氧化还原条件等古环境要素[1,2,3,4]。因此,认识这些要素对于明确烃源岩形成与分布具有重要意义。然而,随着油气勘探不断向深层和超深层领域拓展,古老烃源岩越来越受到关注的同时,其年代老、埋藏深、经历热演化程度高的特性也导致生物标志物及干酪根显微组分等反映成烃母质信息及沉积环境的常规地球化学指标失去指示意义[4]。氮(N)是许多重要的有机组分如蛋白质、核酸和氨基酸等的重要组成元素[5],但是表层海洋生物可利用氮 (大部分为NO3 -)含量很低,它的有限供应成为海洋生态系统营养物质的重要控制因素[6],从而影响着海洋初级生产力。据统计,全球海洋一半以上的原始生产力都受生物可利用氮的控制[7]。原始生产力与有机质的埋藏又会进一步影响海洋水体氧化还原环境[8]。另外,氮也对全球CO2的固定和大气CO2浓度的调节具有重要作用[9],从而影响全球气候变化[10]。因此,对海洋氮循环的研究,有助于深入认识控制烃源岩形成的古环境要素。近年来,氮同位素在反映古环境性质和演化信息上显示出了稳定性,较好地保存了原始地球化学信息,已成为评价海洋氮循环的重要工具[8,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25]。本文系统总结了海洋中的氮循环过程及氮同位素分馏效应,综合不同地质历史时期及现代海洋沉积记录中的氮同位素分布特征,概括了可能涉及的氮循环机制,并对沉积岩氮同位素组成在古环境恢复及其在油气领域应用的现状进行了综述,以提升对氮同位素数据分析及其应用的认识。
1 氮的海洋生物地球化学循环
海洋中氮的溶解态形式主要有以下几种,包括硝酸盐(NO3 -)、亚硝酸盐(NO2 -)和铵(NH4 +)。这些能够被浮游生物直接吸收的活性氮的最终来源为大气N2,通过不同氮库之间的转换,构成了大气—海洋氮循环过程。
由于固氮作用是一个相对消耗能量的过程,因此只有当海洋中可被生物利用的氮被完全消耗时,固氮作用才会发生。
2 氮同位素分馏与氮同位素组成的分布及其控制因素
2.1 海洋氮循环过程中的氮同位素分馏
氮同位素组成是研究海洋中氮的内部循环及输入/输出情况的重要手段,有助于了解海洋氮循环及其对海洋主要氮库的同位素组成产生的影响[31,34,35,36,37]。同位素分馏一般分为平衡分馏和动力分馏。海洋中氮同位素的变化通常以动力分馏为主,发生于从一种形式的氮向另一种形式的氮的转化过程。动力分馏因子ε的定义是在某一反应中2种同位素由反应物向产物转化的速率差异。比如,某一反应的ε为5‰,那么任何时候产物的氮同位素组成始终要比反应物的氮同位素组成低~5‰。ε(‰)=(14 K/15 K-1)×1 000。其中,14 K和15 K分别是含有14N和15N反应物的速率系数。当ε≤1 000‰时,ε约等于反应物与其相应产物的氮同位素组成之差(ε≈δ15Nreact-δ15Nproduct)。海洋沉积物的氮同位素组成取决于海水中生物可利用氮的同位素组成,以及在同化过程及随后在水体和沉积物中发生的一系列转化过程发生的分馏[24]。
2.1.1 固氮作用
表1 氮循环过程中发生的分馏情况(分馏因子ε≈δ15Nreact-δ15Nproduct) (据文献[22,45],修改)Table 1 Isotope fractionations in the process of nitrogen cycle (fractionation factor ε≈δ15Nreact-δ15Nproduct) (revised according to Refs.[22,45]) |
| 氮循环过程 | 同位素分馏范围/‰ | 保存情况 |
|---|---|---|
| 固氮作用 | Mo固氮酶 -2~+1 | 保存在氮受限的生态系统中的沉积物内 |
| Fe-V固氮酶 +6~+8 | ||
| 铵同化作用 | +14~+27 | 无记录 |
| 硝化作用 (NH4 +→NO2 -) | +14~+38 | 在化跃层进行完全,无保存 |
| 硝化作用 (NO2 -→NO3 -) | -12.8 | 在化跃层进行完全,无保存 |
| 硝酸盐同化作用 | +5~+10 | 在透光带进行完全,无保存 |
| 异化硝酸盐还原作用 | +30? | — |
| 反硝化作用 | +10~+30 | 通常在次氧带发生不完全反硝化,保存在同化NO3 -的生物体中 |
| 厌氧氨氧化作用 | +16或+23~+29 | 很难与反硝化区分开 |
2.1.2 再矿化作用
2.2.3 同化作用
2.1.4 硝化作用
2.1.5 异化硝酸盐还原作用(DNRA)
2.1.6 反硝化作用与厌氧氨氧化作用
海洋中可被生物利用的氮通过反硝化及厌氧氨氧化移除,14NO3 -会被优先利用,NO3 -库中会相对富集较重的15N[31]。若反硝化作用过程发生在水体环境中,特别是在最小含氧带(OMZ),氮的氧化物形式(NO3 -及NO2 -)向气态的N2O或N2的转化不完全,会使残余NO3 -库明显富15N,从而产生较大的同位素分馏(ε≈+20‰~+30‰)[52,53](表1)。但是,若反硝化作用过程发生在次氧环境的沉积物中,那么这一过程会将孔隙水中的NO3 -近乎消耗完全,一般产生的分馏很小(ε≈+1‰~+3‰)[36,54]。在厌氧氨氧化作用过程中,NH4 +被NO2 -氧化为N2 [55],这一过程产生的同位素分馏并不十分清楚[56],尽管有人曾提出这一过程产生的同位素分馏可能与反硝化过程相似[28] (表1)。
2.2 沉积岩中氮同位素组成的分布特征及其控制因素分析
2.2.1 偏负的δ15N值
表2 沉积物中氮同位素记录及可能的氮循环作用过程Table 2 Nitrogen isotope records in sediments and possible nitrogen cycling processes |
| δ15N分布 | 水体中可能占主导的N循环 | 沉积物δ15N值/‰ | 地区 |
|---|---|---|---|
| <-2‰ | 真光带缺氧区绿硫或紫硫 细菌同化作用 | 0~-8 | 现代黑海[60]、东地中海[61] |
| -4.7 | 华南三岔剖面寒武系牛蹄塘组[76] | ||
| 不完全铵同化作用 | <-2 | 印度古元古代晚期Aravalli Supergroup[59] | |
| ~-7.5 | 华南地区寒武系(2~3阶)[44] | ||
| -2‰~+1‰ | 蓝细菌固氮作用 | 0~+2 | 华南三岔剖面寒武系牛蹄塘组[76] |
| -2~+3.1,均值0±1 | 华南Chunye1井寒武系荷塘组[72] | ||
| ~-2 | Cariaco盆地[77] | ||
| -2~+1 | 黑海[64] | ||
| -1~+1 | 华南南盘江盆地太平剖面上二叠统—下三叠统[78] | ||
| >+2‰ | 不完全硝化作用 | +2~+18 | 白令海[66,67] |
| 不完全反硝化作用 | -2.6~+4.8,均值2.2±1.3 | 华南Chunye1井寒武系皮园村—荷塘组[72] | |
| +4~+9 | 华南毛石—中南剖面震旦系灯影组[73] | ||
| +4~+8 | 华南三峡地区震旦系陡山沱组[71] | ||
| +7.4 | Santa Barbara盆地[69] | ||
| >+11 | 阿拉伯海[70] | ||
| 硝酸盐同化、固氮及反硝化作用相平衡 | +1~+16,均值~+5 | 现代海洋沉积物[75] |
2.2.2 接近0‰的δ15N值
2.2.3 偏正的δ15N值
海洋氮循环中能够使δ15N值偏正的过程主要有不完全同化作用、不完全硝化作用和不完全反硝化作用。尽管不完全同化作用也可以使残余氮库富集重的氮同位素,但是关于这一作用的地质记录鲜有报道[64]。不完全同化作用会导致水体中残余的NH4 +库富15N[46],通过同化作用使生物体或沉积物中的δ15N呈正值[65]。由于硝化作用进行的速度太快[60],所以一般情况下进行的相对完全。目前仅在白令海地区有关于不完全硝化作用的报道,沉积物中有机质的δ15N值分布在+2‰~+18‰之间[66,67](表2)。不完全反硝化作用主要发生在氧化/次氧化界面,是导致现代海洋NO3 -氮同位素组成变重的主要原因,这些NO3 -会被生物体同化并将富15N的特征记录下来[31,37]。而且不完全反硝化作用越强,δ15N值可能会越高[68]。比如Santa Barbara盆地及阿拉伯海次氧环境下发生了强烈的不完全反硝化作用,沉积物的δ15N值分别为~+7.4‰[69]及>+11‰[70]。另外,古代沉积物中也有关于不完全反硝化作用的记录,如华南地区寒武系、震旦系地层中高达~+11‰的δ15N值[44,71,72]。
3 沉积岩氮同位素数据的可靠性分析
成岩及变质作用会对海洋沉积物中有机质的δ15N值产生影响。在早期成岩过程中,氮会在微生物的脱氨基作用下发生丢失,可能会释放较轻的NH4 +进入沉积物孔隙水[79],导致残余有机物富集15N,同位素值会增加千分之几[80,81]。如果这些NH4 +被黏土矿物捕获,那么全岩沉积物的δ15N值发生微弱变化或不发生变化[58]。随着埋藏温度的升高,有机分子可能会进一步降解,产生难降解的有机质[82]。这可能会导致氮进一步释放到流体中,随后被黏土捕获或凝聚成更复杂的有机分子。研究表明,在缺氧水柱的早期成岩过程中,有机质的δ15N值通常变化不大或略有下降(降低~1‰)[83,84]。但在有氧条件下,δ15N值可能会增加3‰~5‰[24,85]。
成岩作用后,变质过程会进一步影响氮同位素组成。在这一过程中,14N相对15N会被优先移除沉积物[86],因此,一般会造成N同位素组成变重及全岩N同位素与干酪根N同位素之间的分异[23]。氮同位素值的变化取决于变质级别:对于角闪岩相,δ15N值会增加3‰~4‰[25,87];但在绿片岩相及亚绿片岩相中,δ15N值会分别增加1‰~2‰和<1‰[88]。若地层中δ15Nsed高于δ15Nkerogen,则推断沉积岩经历了变质级别相对低的绿片岩相[89]或者是级别更高的变质过程[90],若二者值相近,则可认为沉积物保存了原始的δ15N特征[58,91]。因此,有观点认为对于变质级别低于绿片岩相的沉积岩,氮同位素组成的原始特征不会发生显著变化,相应的δ15Nsed数据更可靠[11]。
判断成岩及变质作用是否对原始氮同位素组成产生影响,通常利用总有机碳(TOC)、总氮(TN)、有机碳同位素(δ13Corg)及δ15N之间的相关关系进行判断。现代海洋有机质的C/N值为~6.6[92],而持续的变质作用会导致N相对C的优先损失,使C/N值增加至数百或上千[86]。但此时,如果TN与TOC总体表现线性关系,则说明氮来源于原始有机质[93],而TN轴的截距则反映的是无机黏土束缚的氮组分,很可能来自于微生物对有机质的硝化作用[79]。另外,由于成岩或变质过程中的热演化作用会优先将14N移除沉积物,因此,若δ13Corg与δ15N正相关,TN与δ15N、δ15N与C/N值负相关,则说明全岩氮同位素遭受沉积后作用的影响而发生改变[15,23];若δ15N与TOC,δ15N与TN及δ15N与C/N值之间没有线性关系,则表明原始氮同位素组成在成岩过程中未发生明显变化[76]。
4 氮同位素在烃源岩古环境恢复中的应用
4.1 氮同位素对古氧化还原环境的指示
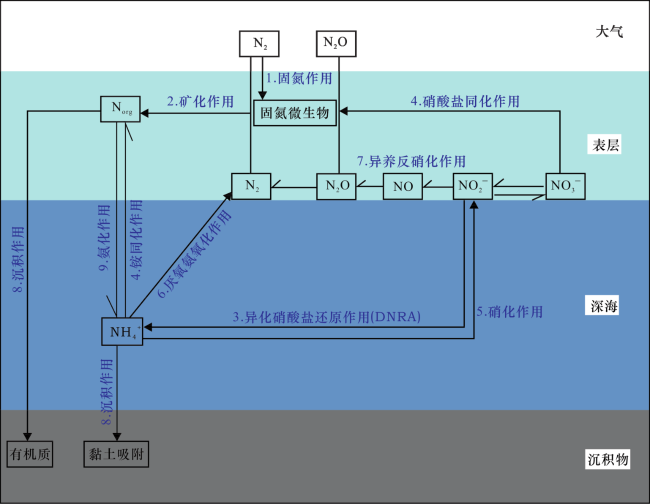
在含氧的海洋环境中,如现代海洋[图2(a)],生物可利用氮的主要形式为NO3 -,它是由NH4 +经硝化作用而来的。表层水体透光带的NO3 -和NH4 +被浮游生物大量同化,几乎不产生分馏,而深处的NO3 -随着上升流经过最小含氧带(OMZ)时,会发生反硝化,导致残余NO3 -库富集15N。在这种水体环境下,NO3 -的损失与N2的固定主要取决于大洋环流模式、深水与中等深度水体的氧化还原条件以及上部沉积物氧化还原层的深度[43],这时氮同位素组成的分布特征应该代表海洋氧化的程度[96]。现今海洋表层沉积物的δ15N值主体分布在5‰~6%之间,最大值可达~+15‰[75]。早古生代和中生代海洋可能具有类似的水体结构[11],但是海洋中含氧量要远较现代海洋低[图2(b)],导致最小含氧带(OMZ)的NO3 -近乎完全反应,从而导致δ15N值降低[12]。
在氧化还原分层的海洋中,如古代的元古宙、显生宙海洋及现代的黑海、Cariaco盆地[97,98,99,100,101],NO3 -主要分布于氧化还原过渡带之上,而NH4 +位于过渡带之下的缺氧水体中。对于氧化还原过渡带相对稳定的现代海洋环境[图2(c)],表层水体中的NO3 -一方面被同化作用消耗,另一方面在氧化还原过渡带附近发生反硝化,而缺氧水体中的NH4 +在过渡带会经历硝化—异养反硝化或厌氧氨氧化作用反应完全,转化为N2或N2O[60,101]。这会引起生物可利用的氮发生大量损失,而使固氮作用占主导,导致δ15N接近于0‰[96]。对于存在上升流的具有不稳定氧化还原过渡带的分层水体,如果过渡带较深[图2(d)],上升流携带的NH4 +在被完全转化为N2或N2O之前,会有部分穿越过渡带,使富15N的NH4 +可以到达透光带,从而被直接同化或被氧化为NO3 -后发生同化,产生偏正的δ15N值[59,90]。相比之下,如果氧化还原过渡带较浅[图2(e)],上升流可能会直接进入海洋表层[102],携带的NH4 +会被完全同化,并进一步被硝化—异养反硝化及厌氧氨氧化作用转化为N2或N2O。若NH4 +同化作用占主导,则会导致δ15N值偏负[58],而当N2或N2O转化作用占主导时,则会使δ15N趋于正值[59,103,104]。
4.2 氮同位素分布对生物演化的响应
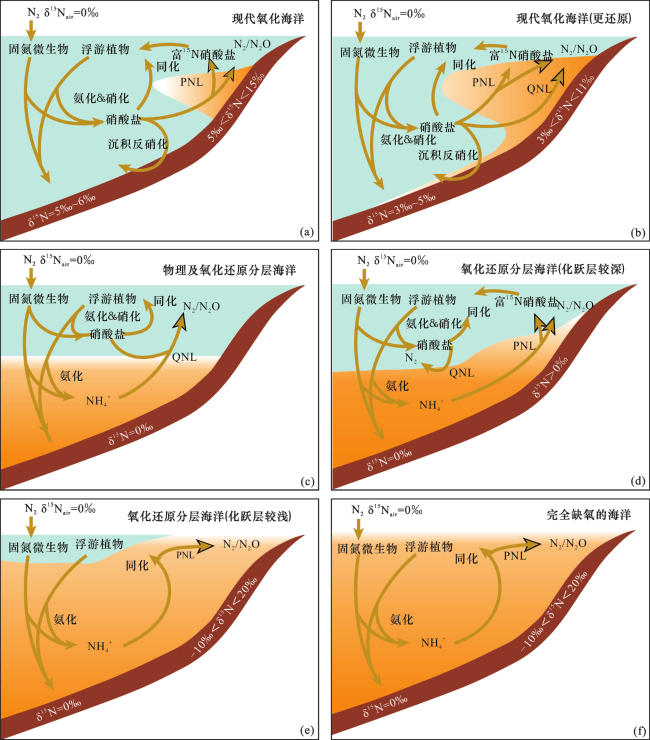
在大成氧事件(GOE, 2.4~2.0 Ga)之前,人们认为海洋磷元素的稀缺限制了初级生产力,直到古元古代海洋溶解氧和硫酸盐含量上升[112],海洋碳循环的速度和初级生产力才可能随着地球表面环境的氧化而增加[113]。在GOE期间,磷含量的增加[113]可能导致氮成为海洋环境中的限制性营养元素。然而,古元古代海相沉积岩中的氮同位素组成分布 (图3)并未显示出这一时期存在氮限制作用的证据[15]。恰恰相反,古元古代海相页岩δ15N值的持续升高表明,海洋中生物可利用的氮是足够的,可以为初级生产提供动力。尽管关于真核生物出现的最早时间仍存在争议,可能在大约1.7 Ga左右[114]或者更早[115],但是,已有研究表明具有甾醇合成性能(真核生物的一个特性)的生物体可能在GOE开始时已经开始进化了[116]。因此,古元古代δ15N高值可能指示了当时海洋中存在具有NO3 -同化能力的生物,虽然无法确定这些生物是否是真核生物,但至少说明当时海洋中存在足够的生物可利用的NO3 -,从而缓解了真核生物繁殖面临的氮元素限制。
在新元古代的埃迪卡拉纪,氮同位素组成数据似乎也表明海洋中存在有氧氮循环 (图3),使得表层水体NO3 -含量丰富。丰富的NO3 -可能有助于埃迪卡拉纪宏观多细胞藻类的进化[117]。成冰纪冰消期之后的风化作用增强[118],限制生物生长的营养物质(磷和氮)的供应增加改变了初级生产者的生态结构,可能促进了浮游植物结构的转变。从生物标志物等数据[119]可以看出,在新元古代,浮游植物由小细胞的蓝藻向大细胞的浮游藻类转变。这与氮同位素数据所揭示的结果相同,即在埃迪卡拉纪晚期海洋中存在一个稳定的NO3 -库,真核浮游植物会优先吸收NO3 -,营养氮元素的增加可能促进了真核浮游植物向大细胞演化,并可能显著改变了生物泵的架构,从而使更多的有机氮在海洋深处富集[44]。
4.3 氮同位素分布与气候环境变化的关系
氮对海洋生产力和有机碳通量起重要控制作用[122],因此潜在地对全球气候系统产生重要影响[10]。反硝化过程中NO3 -的还原需要NO2 -还原酶和NO还原酶等进行催化,其中间产物N2O为温室气体。在地质历史时期,大气N2O含量往往在冰期较低,而在间冰期明显升高。这表明反硝化过程可能通过释放N2O而影响全球气候变化[10]。比如,第四纪(2.6 Ma至今)海洋沉积物氮同位素组成的变化被认为与更新世冰期—间冰期有机氮埋藏及海洋反硝化速率的变化有关[68,77,123]。海洋氮循环对气候可能会产生积极的反馈作用,尽管有人提出固氮及反硝化2种影响海洋氮库的关键性氮循环对气候起到了负反馈作用[35]。LUO等[78]对华南南盘江盆地二叠纪—三叠纪过渡期2个含微生物岩的剖面开展了高分辨率全岩有机碳(δ13Corg)和氮(δ15N)同位素分析。研究发现2个剖面的δ15N值在二叠纪生物大灭绝后立即发生负偏移,从+3‰变化为0‰,同时δ13Corg也发生了明显的负偏移。δ15N的持续低值表明,在大灭绝后,微生物固氮作用成为海洋生物可利用氮的主要来源,而固氮作用的增强可能预示着当时以强烈反硝化和/或厌氧氨氧化为特征的分层缺氧水体的盛行。此外,温室气体N2O的释放,可能进一步导致海洋生物灭绝之后气温升高。
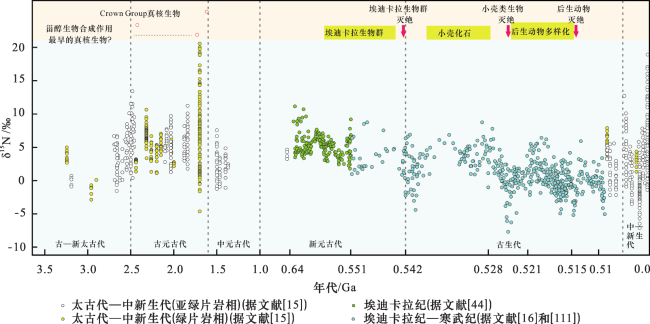
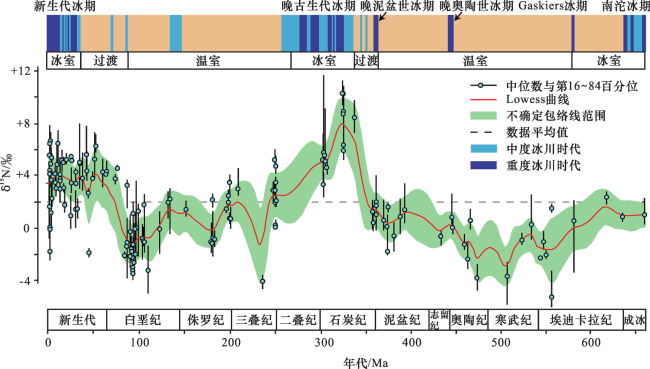
同样地,ALGEO等[124]对660 Ma以来海相沉积单元的氮同位素组成研究也表明,海洋氮循环变化与气候变化之间存在协同关系。在显生宙,温室(greenhouse)气候模式下的δ15N值较低(约为-2‰~ +2‰),而冰室(icehouse)气候模式下的δ15N值较高(约为+4‰~+8‰)(图4)。在冰室早期,δ15N迅速变高,表明海洋氮循环可能对长期气候变化产生重要作用。尽管海洋氮循环会驱动气候发生长期变化的观点是推测的,但也不能完全否定。海洋氮循环与海洋有机碳的埋藏密切相关[125,126],但是它是对碳通量变化的被动响应,因此,氮循环可能在气候变化过程中起到了重要的放大器作用。例如,在气候变暖的较长时间间隔内,海洋低氧区域N2O产量的增加可能会起到积极的气候反馈作用[127],从而促进发育长期的气候双峰模式(即温室与冰室模式)。
5 氮同位素在油源对比中的应用
除了在古环境恢复方面发挥的重要作用,氮同位素在油气领域,如原油类型识别及油源对比等方面也呈现出了巨大的应用潜力[128,129,130]。陈传平等[128]认为不同性质原油δ15N值主要分布在-6‰~+20‰之间,其中淡水环境原油δ15N值低,大致在+1‰~ +5‰之间,咸水和半咸水沉积环境形成的干酪根和原油δ15N值较高,通常大于+10‰,甚至超过+17‰。同样地,刘娅昭等[131]对辽河地区陆相及塔里木盆地海相原油氮同位素组成分布的研究表明,海相烃源岩形成的原油氮同位素组成要比陆相烃源岩形成的原油氮同位素组成偏轻。另外,有研究认为,原油的δ15N值基本不受成熟度、运移距离和生物降解作用的影响[132],但是也有观点认为相同沉积环境下,原油受埋深高温裂解作用影响,会使原油中富集14N,导致高成熟度油的氮同位素组成偏轻[131]。因此,尽管原油氮同位素组成有望成为新的油源对比指标,但是其受热演化的分馏效应仍不明朗,需要开展更多的热模拟实验,以探究其在油源对比研究中的适用条件。随着勘探目标逐渐转向深层和超深层,油气性质的复杂多样性、以及多种相态油气共存等现象逐渐突出[133,134,135],对油气成因的判识指标的要求也越来越高。另外,随着页岩气向寒武系、震旦系等古老层系的推进,古老黑色页岩的成因机理将成为研究热点[136,137,138]。因此,氮循环及氮同位素的研究将成为一个新的热点,并有望为这些复杂石油地质难题的解决提供新的手段。
6 总结
对海洋氮循环的研究,有助于深入认识控制烃源岩形成的古环境要素。N同位素对评估古海洋氮循环十分重要,偏负的δ15N值可能与不完全铵同化作用或绿硫/紫硫细菌的同化作用有关,近0‰的δ15N值分布可能与固氮作用有关,而偏正的δ15N值可能反映了不完全同化作用、不完全硝化作用或不完全反硝化作用的影响。
由于氮同位素组成受海洋水体氧化还原条件的影响较大,对于反映局部水体氧化还原特征可能更加灵敏,因此,开展区域性或者全球性海洋生物地球化学研究,需要与其他海洋环境指标如钼同位素及铀同位素等综合使用。
地史时期的氮同位素组成在分布上与生物辐射及灭绝,温室及冰室气候变化呈现出了显著的耦合性,对研究环境演化、生物演化及二者关系具有重要指示作用,对重建烃源岩发育环境、揭示古老烃源岩发育机制具有重要意义。另外,氮同位素在油气领域如原油类型判识及油源对比方面也表现出了巨大的应用潜力。
 甘公网安备 62010202000678号
甘公网安备 62010202000678号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}