

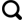
0 引言
表1 生物扰动储层研究实例统计Table 1 Research examples of bioturbated reservoirs |
| 地点/国家 | 地层/ 界面名称 | 地质年代 | 岩性 | 沉积环境 | 物性特征 | 参考文献 |
|---|---|---|---|---|---|---|
| 威拉帕湾,美国 | Glossifungites 固底界面 | 第四纪 | 基质为泥岩,潜穴被砂质充填 | 潮间带 | 潜穴的渗透率显著高于基质 | Gingras等[4] |
| 阿尔伯塔,加拿大 | Palliser组 | 泥盆纪 | 潜穴为白云岩,基质为灰岩 | 浅水碳酸盐岩缓坡 | 潜穴的渗透率高于基质 | Gingras等[5] |
| 挪威海 | Lysing组 | 白垩纪 | 细砂岩、粉砂岩、泥岩 | 大陆架—外陆架 | 潜穴的渗透率高达基质的2个数量级 | Gingras等[6] |
| 加瓦尔油田, 沙特阿拉伯 | Glossifungites 固底界面 | 侏罗纪 | 基质为泥晶灰岩,潜穴为砂糖状白云岩 | 滨岸 | 潜穴区域为高渗区 | Pemberton等[7] |
| 东爪哇海 | Paciran组 | 中新世 | 砂岩、灰岩 | 深海远洋 | 潜穴间的连通性好, 渗透率高于基质 | Pemberton等[7];Noble等[8] |
| 挪威近海 | Ula组 | 侏罗纪 | 砂岩 | 滨岸 | 强烈的生物扰动提高潜穴的孔隙度5%~10% | Gingras等[6] |
| 纽芬兰,加拿大 | Ben Nevis组 | 白垩纪 | 砂岩 | 海湾 | 生物扰动使得潜穴的孔渗提高6倍或降低1/3 | Tonkin等[9] |
| 阿尔伯塔,加拿大 | Alderson组 | 白垩纪 | 含薄泥质夹层的细砂岩 | 三角洲海岸 | 潜穴的渗透率显著增高 | Hovikoski等[10] Lemiski等[11] |
| 阿尔伯塔,加拿大 | Bluesky组 | 白垩纪 | 隐蔽生物扰动砂岩 | 滨岸 | 孔隙度、渗透率略有改善且数值更为集中 | Gordon等[12] |
| 曼尼托巴,加拿大 | Red River组 | 奥陶纪 | 斑状白云化灰岩 | 陆缘碳酸盐岩台地 | 基质的平均渗透率为1.65×10-3μm2,潜穴为19.2×10-3μm2 | Gingras等[5] |
| 西北地区,加拿大 | Lonely Bay组 | 泥盆纪 | 潜穴发生钙质胶结 | 滨岸/远岸 | 潜穴部分晶内孔发育,孔隙度增加 | Corlett等[15] |
| 塔里木盆地,中国 | 东河塘组 | 泥盆纪 | 砂岩 | 潮汐砂坝 | 储层物性随生物扰动增强而变差 | 齐永安[16] |
| 东濮凹陷,中国 | 沙四段 | 古近纪 | 粉砂岩 | 滨浅湖—深湖 | 生物扰动部位的泥质含量低,胶结物含量高 | 纪友亮[17] |
1 地质背景

2 样品与方法
3 Kh2段遗迹学与非均质性特征
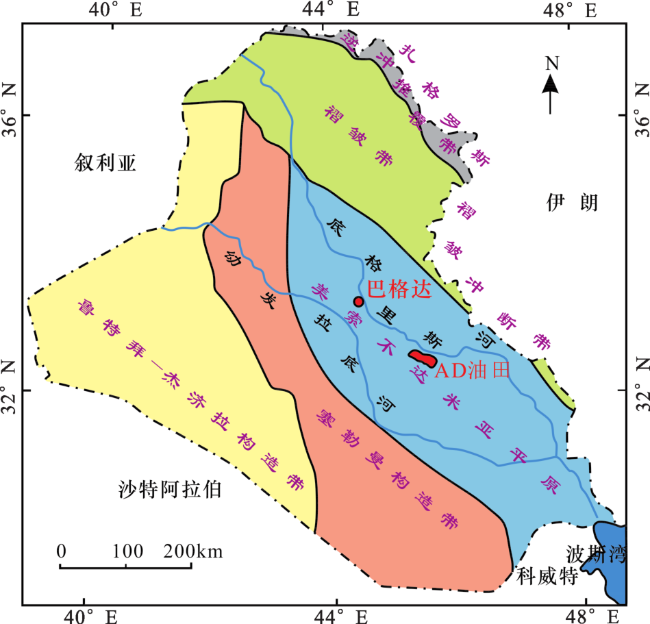
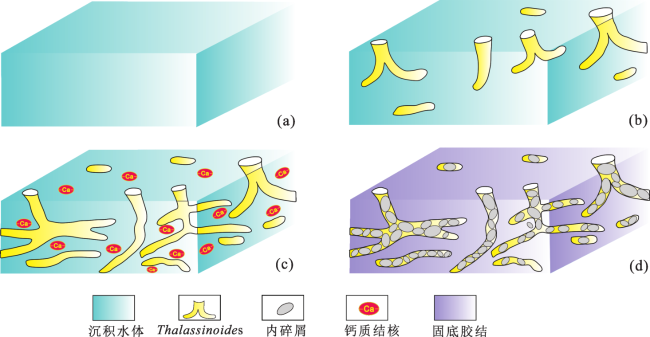
3.1 遗迹学特征

图3 AD油田Kh2段典型遗迹学特征(a) Thalassinoides, A3井, 2 674.8m, MFT3, Kh2-3; (b) Thalassinoides, A3井, 2 673.6m, MFT3, Kh2-3; (c) Rhizocorallium, A2井, 2 662.0m, MFT1, Kh2-5; (d) Planolites, A4井, 2 629.30m, MFT1, Kh2-4; (e) Ophiomorpha, A1井, 2 649.5m, MFT4, Kh2-2;(f) Ophiomorpha, A2井, 2 666.8m, MFT4, Kh2-2; (g) Ophiomorpha, A3井, 2 662.78m, MFT4, Kh2-1-2U;(h) Ophiomorpha, A3井, 2 663.2m, MFT4, Kh2-1-2U; (i) Thalassinoides, A1井, 2 648.5m, MFT5, Kh2-1-2L;(j) Thalassinoides, A3井, 2 664.1m, MFT5, Kh2-1-2L; (k)、(l) Glossifungites, A4井, 2 609.2m, MFT5, Kh2-1-2L Fig.3 Diagram of typical ichnology characteristics of Kh2 member in AD Oilfield |
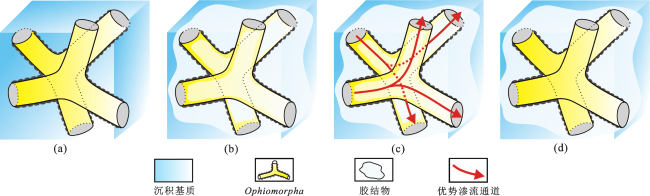
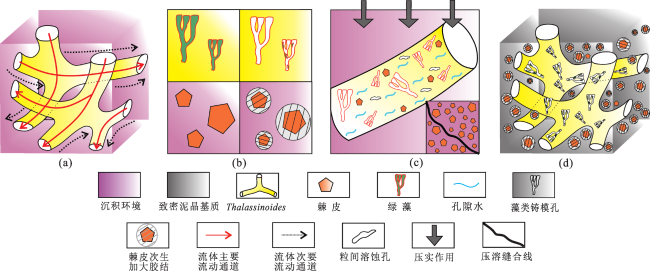
3.2 储层非均质性特征
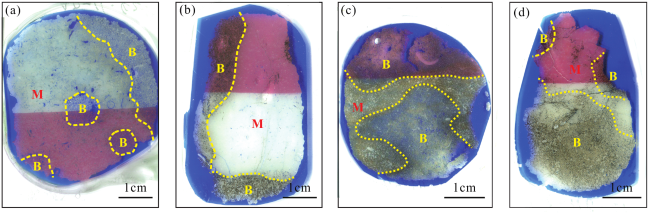

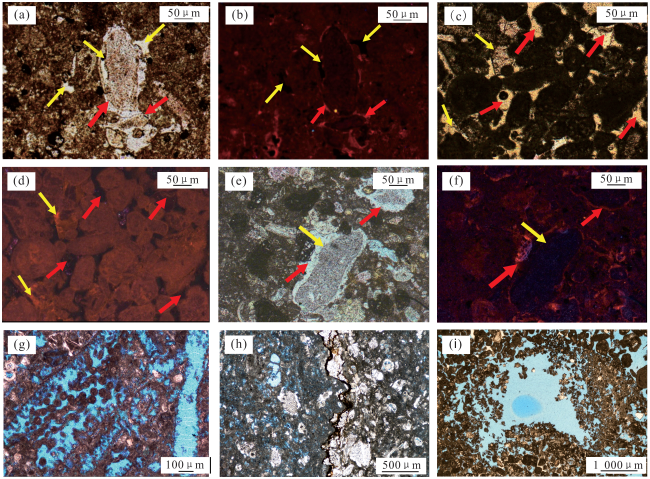
图6 Kh2段松散底质颗粒滩储层的非均质性特征(a)—(f) 生物扰动及孔隙发育特征显微镜下照片;(a)—(c) A2井,2 636.7m, Kh2-1-2L;(a) 潜穴与基质交界,单偏光;(b) 基质,单偏光;(c) 潜穴,单偏光;(d)—(f) A3井, 2 665.4m, Kh2-1-2L;(d) 潜穴与基质交界,单偏光;(e) 基质,单偏光;(f) 潜穴,单偏光;(g) 不同含油性分布特征素描图,Ophiomorpha潜穴及其周缘的含油性较好,Kh2-1-2U中下部;(h) 点渗透率测试,Ophiomorpha遗迹组构,MFT4,A1井2 649~2 650m,Kh2-1-2L,红蓝色充填圆圈代表探针实测位置,红色对应渗透率高值,蓝色对应渗透率中—低值,渗透率单位为×10-3µm2 Fig.6 Heterogeneity characteristics of Kh2 bioturbated hardground grain beach reservoir |

图7 Kh2段固底底质颗粒滩储层的非均质性特征(a)—(d) 生物扰动及孔隙发育特征显微镜下照片,A3井, 2 664.1m, Kh2-1-2U; (a)、(b) 潜穴与基质交界,单偏光;(c) 潜穴,单偏光;(d) 基质,单偏光;(e) 不同含油区域分布特征素描图,Thalassinoides潜穴及周缘的含油性显著优于受强烈胶结改造的基质,Kh2-1-2U中下部;(f) 点渗透率测试,MFT5,A2井,2 638~2 639m,Kh2-1-2L,渗透率单位为×10-3µm2 Fig.7 Heterogeneity characteristics of Kh2 bioturbated looseground grain beach reservoir |

图8 Kh2段绿藻滩储层的非均质性特征(a)—(d) 生物扰动及孔隙发育特征显微镜下照片,A1井, 2 657.5m, Kh2-3; (a)、(b) 潜穴与基质交界,单偏光;(c) 基质,单偏光;(d) 潜穴,单偏光;(e) 不同含油区域分布特征素描图,Thalassinoides潜穴周缘的含油性优于基质,Kh2-3;(f) 点渗透率测试,MFT3,A1井,2 657~2 658m,Kh2-3,渗透率单位为×10-3µm2 Fig.8 Heterogeneity characteristics of Kh2 bioturbated green algae reservoir |
 甘公网安备 62010202000678号
甘公网安备 62010202000678号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}